Dýchací systém
Hlavním orgánem dýchacího (pulmonálního) systému jsou plíce. Vzduch mezi vnějším prostředím a plicemi proudí v dýchacích cestách, které tvoří: dutina nosní a ústní, nosohltan, hrtan, průdušnice a průdušky, které ústí do plicních laloků. V nich se z průdušek stávají průdušinky, které slouží k přenosu vzduchu do tenkostěnných plicních sklípků (alveolů). Hlavním dýchacím svalem je bránice, které v mechanické práci napomáhají ještě pomocné nádechové a výdechové svaly.
Hlavní funkcí dýchacího systému je společně s kardiovaskulárním systémem zabezpečit dodávku kyslíku a odvod CO2 do/z pracujících svalů.
Regulace plicní ventilace je zprostředkována odpovědí centrálních a periferních chemoreceptorů, které informují respirační centrum uložené v prodloužené míše o změnách parciálních tlaků (p) CO2 a kyslíku (pO2). Periferní chemoreceptory reprezentují karotická a aortální tělíska, která jsou citlivá primárně na změny v pO2, ale částečně reagují také na vzestup pCO2 a H+. Naopak, centrální chemoreceptory, které jsou uloženy v prodloužené míše, stimuluje zejména zvýšení pCO2 a H+ v cirkulující krvi.
Celý proces dýchání má tři mezistupně: 1. výměna plynů mezi okolním prostředím a plícemi (plicní ventilace); dále 2. výměna plynů mezi alveoly a kapilární krví na alveokapilární membráně, jejímž principem je rozdílná hodnota pO2 a pCO2 před (prostor plic) a za membránou (kapilární řečiště). Při nádechu je do plic nasáván vzduch o vyšším pO2 než je jeho hodnota v kapilární krvi. Proto O2 na základě tlakového gradientu (difuze) prostupuje membránou z plic směrem do krve. Stejný princip funguje při výměně CO2, jenom v opačném směru. V krvi jsou oba plyny vratně (reverzibilně) navázány na základě parciálních tlaků na erytrocyty, respektive na hemoglobin (červené barvivo) a konkrétně na část zvanou hem, kde se nacházejí čtyři dvojmocné ionty železa (Fe2+). Posledním, třetím, mezistupněm přenosu plynů je výměna O2 a CO2 mezi krví a svalovou buňkou, který se nazývá respirace. Dalším termínem spojeným s transportem dýchacích plynů v krvi je saturace, která vyjadřuje procentuální podíl nasycení arteriální krve kyslíkem. Změny vazby O2 na hemoglobin se dají vyjádřit pomocí tzv. disociační křivky. Posun disociační křivky směrem doprava vyvolávají následující faktory: a) ↓pH (↑koncentrace H+), b)↑ teplota, c) ↑pCO2,d) ↑koncentrace 2,3 difosfoglycerátu (produkt štěpení glukózy). Pravostranný posun znamená nižší afinitu hemoglobinu na kyslík a snadnější vyvázání O2 z vazby na hemoglobin.
Na začátku zatížení dochází vlivem rostoucí sympatoadrenální aktivity ke zvýšení minutové plicní ventilace (V), která se vyjadřuje v litrech/minutu. Ventilace je součinem dechového objemu (Tidal Volume – TV), který je v klidu u zdravého člověka kolem půl litru, a dechové frekvence (DF), která se v klidu pohybuje v rozmezí od 12 do 16 dechů. Čili ventilace v klidových podmínkách činí kolem 6–8 L/min. Z proventilovaných 6 L vzduchu v organismu zůstávají asi 4 % kyslíku. Poměr ventilace a spotřeby kyslíku (V/VO2) se označuje jako ventilační ekvivalent pro kyslík. Jeho hodnota udává kolik množství vzduchu je potřeba za minutu proventilovat, abychom získali jeden litr kyslíku. Na základě tohoto parametru lze posuzovat účinnost (efektivitu) ventilace. U zdravého člověka se tato hodnota pohybuje kolem 24. Vlivem zvyšujícího zatížení roste poptávka pracujících svalů po dodávce kyslíku a odvodu CO2 což vede ke zvýšení plicní ventilace. Na jejím vzestupu se podílí jak prohloubené dýchání (↑DV), tak především ↑DF. Po dosažení úrovně anaerobního prahu, dochází vlivem zvýšeného příspěvku anaerobního štěpení sacharidů na resyntéze ATP k vzestupu koncentrace iontů vodíku (H+). Jejich zvýšená hladina způsobuje pokles pH (acidózu) a tím narušení acidobazické rovnováhy. Výsledkem regulačních mechanismů udržujících acidobazickou rovnováhu, které představují pufrovací (nárazníkové) systémy, je zvýšení pCO2 v krvi (obrázek 1.6). Vzestup pCO2 podráždí centrální chemoreceptory, které vysílají impulz do dýchacího centra s cílem zvýšit plicní ventilace (hyperventilace), na které se nejvíce podílí vzestup DF. Disproporční zvyšování ventilace vůči spotřebě kyslíku a intenzitě zatížení vede k růstu ventilačního ekvivalentu pro kyslík, který v maximu dosahuje hodnoty i přes 35 L, což svědčí o zhoršování účinnosti dýchání při náročné svalové práci. Při maximálním zatížení se uvádějí hodnoty plicní ventilace u sportovců kolem 180 až 200 L při DF 60 a zhruba 60 % TV, což může odpovídat u dospělého muže zhruba 3 L.
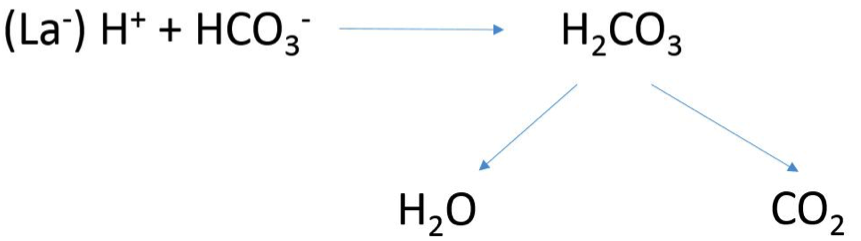
Obrázek 1.6 Regulace acidobazické rovnováhy pomocí uhličitanového pufru
Kardiovaskulární systém
Kardiovaskulární systém (KVS) se skládá ze srdce a cév (tepen a žil), ve kterých jako v uzavřeném systému cirkuluje transportní médium – krev. Hlavním úkolem KVS je transport kyslíku z plic ke svalovým buňkám a odvod CO2, z plodin metabolismu včetně vznikajícího tepla ze svalů.
U srdce rozlišujeme čtyři hlavní části – dvě předsíně a dvě komory (pravou a levou), které jsou propojeny chlopněmi. U srdeční práce, někdy nazývané také jako srdeční revoluce, rozlišujeme – systolu (stažení), kdy dochází k vypuzení krve ze srdce do cév a diastolu (ochabnutí), kdy naopak je srdce plněno krví, kterou do pravého srdce přivádí horní a dolní dutá žíla. Za hlavní regulátor srdeční činnosti je považována aktivita autonomního nervového systému, která srdce ovlivňuje prostřednictvím dvou větví – sympatiku (stimulační efekt) a parasympatiku (relaxační efekt). Kromě regulační aktivity ANS se na řízení srdeční činnosti uplatňují i stresové hormony, zejména adrenalin. Za fyziologických podmínek je sinoatriální uzel klíčovým regulačním místem, kde vzniká akční potenciál, který se dále šíří převodní soustavou srdeční (princip vše nebo nic) a dochází ke kontrakci myokardu (systole). Hlavní funkcí srdce je přečerpávat krev. Srdce společně s cévním systémem tvoří dva oběhy nazývané jako malý (plicní) a velký (tělní). V malém plicním oběhu (pravá komora – plicnice – plíce – čtyři plicní žíly – levá síň) dochází k okysličení krve a odvodu CO2. Okysličená krev se vrací zpět do srdce, kde je dále přečerpávána do velkého tělního oběhu (levá předsíň – levá komora – aorta), který začíná hlavní tepnou – aortou. Zpět do srdce je krev z celého těle přiváděna pomocí žilního systému (horní a dolní dutá žíla – pravá předsíň – pravá komora). Krev, které má dospělý muž asi 5 L, se dělí na formované krevní elementy (erytrocyty, leukocyty a trombocyty) a čirou nažloutlou tekutinu – krevní plazmu. Krev plní funkci a) transportní; b) imunitní; a c) termoregulační.
Při zvýšené potřebě kyslíku pracujícími svaly musí začít srdce přečerpávat větší objem krve. Množství krve, které srdce přečerpá za jednu minutu, se nazývá minutový srdeční výdej (MSV) a za klidových podmínek je rovno asi 5 L. Hodnota MSV je součinem systolického objemu, který je definován jako množství krve, které srdce vypudí při jednom stahu a srdeční frekvence (SF). Hodnotu MSV lze jednoduše vypočítat jako součin SV (70 mL) a SF (70 tep/min). Při postupném zvyšování intenzity zatížení jsou požadavky na rostoucí MSV kompenzovány zvyšujícím se SV (výsledek roztažení myokardu) společně se zvyšováním SF. U zdravého muže ve věku 20–30 let se přibližně od úrovně 100 až 120 tepů (40 % VO2max) na dalším zvyšování MSV začíná výhradně podílet pouze vzestup SF. Od této intenzity dochází k lineárnímu zvyšování SF s rostoucím zatížením, přičemž porušení linearity nastává při intenzitě kolem 170–180 tepů (70–80 % VO2max), kde se nachází anaerobní práh (cirkulační). Při maximálním zatížení může hodnota SF u mladých netrénovaných osob dosáhnout více jak 200 tepů/min, což je při SV = 100 ml množství přečerpané krve odpovídající 20 L/min. Hodnoty SF v podmínkách maximální práce jsou individuálně variabilní, ale orientačně je lze vypočítat ze vzorce 220 – věk. U tohoto vzorce odhadu SFmax se uvádí, že začíná platit přibližně od 15. roku věku.
Arteriovenózní diference kyslíku
Krev je schopna při plném nasycení hemoglobinu vázat 20 ml kyslíku/100 ml. Během tělesné práce, kdy je do pracujícího svalu přiváděna okysličená (arteriální) krev, svalové buňky využijí pouze část této nabídky. Zbytek kyslíku se bez užitku vrací žilním systémem zpět do srdce. Rozdíl v úrovni nasycené arteriální a venózní krve kyslíkem se nazývá arteriovenózní diference kyslíku (a-v)O2. Její hodnota udává extrakci kyslíku periferními tkáněmi. Za klidových podmínek se hodnota (a-v)O2 pohybuje kolem 5 ml/100 ml krve. Při tělesné práci se její hodnota zvyšuje a při maximální intenzitě může dosahovat až 16–17 ml kyslíku/100 ml krve. U vyspělých vytrvalců lze pozorovat hodnoty i 18 ml kyslíku/100 ml krve. Ani při maximální práci však není dosaženo kompletní extrakce kyslíku z venózní krve z důvodu smíchání venózní krve, která protékala pracujícími svaly s krví, která během zatížení protékala na kyslík méně náročnými tkáněmi a část kyslíku si ponechala.
Reakce transportního systému na dynamickou zátěž
Při přechodu z klidového stavu do zatížení rozeznáváme u transportního systému dvě fáze:
1) iniciální rychlou fázi, která trvá přibližně 30–45 sekund a jsou pro ni typické velké funkční změny v transportním systému;
2) přechodovou fázi, která je pomalejší než iniciální fáze. V přechodové fázi dochází k postupnému přizpůsobení transportního systému požadavkům pracujících svalů. Tato fáze přechází v několika minutách při intenzitě zatížení okolo 60 % VO2max do rovnovážného stavu (steady state).Ten je možné definovat jako stav, při kterém se transportní systém vyrovnal se zvýšenými požadavky pracujících svalů na přísun O2 a energie a odvod metabolitů a CO2. V pravém setrvalém stavu osciluje SF v rozmezí +-4 tepy/min. Navození setrvalého stavu determinuje především intenzita zatížení a platí, že čím je vyšší intenzita zatížení, tím později je setrvalého stavu dosaženo. Při vysoké intenzitě zatížení však setrvalý stav nemusí ani nastat. Za nejvyšší intenzitu zatížení, která umožňuje vznik setrvalého stavu, se považuje zatížení odpovídající anaerobnímu prahu nebo úrovni těsně pod ním. Po začátku tělesné práce se v důsledku funkčních změn v transportním systému zvyšuje VO2 a výdej CO2. Úroveň VO2 od započetí práce roste postupně, nejdříve strměji a před dosažením setrvalého stavu již mírněji. Než je dosaženo setrvalého stavu, není zcela pokryta adekvátní dodávka O2 pro aerobní resyntézu ATP, proto si organismus pro zisk ATP musí vypomoci zapojením anaerobních procesů (anaerobní glykogenolýzou), kdy se štěpí výhradně sacharidy za vzniku laktátu + vodíkových iontů (H+). Chybějící množství O2 od začátku práce po dosažení setrvalého stavu se nazývá kyslíkový deficit. Jeho velikost je úměrná intenzitě zatížení. Nižších hodnot kyslíkového deficitu při stejné relativní intenzitě zatížení dosahují více zdatné (trénovanější) a mladší osoby, zejména děti, u kterých byla prokázána rychlejší periferní cirkulace. Během přechodové fáze rozlišujeme okamžik nazývaný jako mrtvý bod. Ten vzniká v důsledku prozatím nedostatečné dodávky O2 pracujícím svalům, kterou organismus supluje zapojením anaerobního metabolismu, který způsobuje acidózu (↓pH – ↑H+) spojenou s mentálním diskomfortem. Narušení acidobazické rovnováhy vyvolává homeostatickou kompenzační odpověď v podobě pufrovacího (nárazníkového) systému (↑pCO2 v krvi + hyperventilace). Pro okamžik vyrovnání požadovaného množství O2 pro svalovou činnost se používá termín druhý dech, po kterém nastává již zmiňovaný setrvalý stav. Při práci v setrvalém stavu může dojít ke zvýšené VO2, než je aktuální potřeba pracujících svalů z důvodu zmírňování kyslíkového deficitu ještě během pohybové aktivity.
Zotavení a zotavný kyslík
V okamžiku přerušení či ukončení tělesné práce postupně klesají energetické nároky pracujících svalů na klidovou úroveň, ale orgány transportního systému (plíce+srdce) i v momentu přerušení zatížení pracují s jistou setrvačností a do předzátěžového stavu se navracejí pomaleji. Typickým projevem pozvolnějšího zotavení transportního systému je zvýšená pozátěžová ventilace, hodnota SF nebo úroveň VO2, která může dosáhnout podle intenzity zatížení své klidové hladiny až za 24 hodin. Zvýšená VO2 nad aktuální energetické potřeby organismu během zatížení se nazývá zotavný kyslík, který v akutní (rychlé) fázi zotavení slouží pro splácení kyslíkového dluhu a pro utilizaci laktátu vzniklého zejména na začátku, ale také v průběhu intenzivní práce, pokud byla prováděna ve vyšších intenzitách (na nebo nad úrovní anaerobního prahu – ANP). Kyslíkový deficit je zpravidla menší než objem přijatého pozátěžového kyslíku. Laktát vyprodukovaný během svalové práce se využívá jako energetický substrát již během zatížení, proto ne zcela přesně reprezentuje rozsah zapojení anaerobního metabolismu. Zotavný kyslík dále slouží pro úhradu zásob O2 v krvi a ve svalovém myoglobinu, k resyntéze ATP a CP. Větší část kyslíku je využita při přeměně laktátu na glykogen v játrech procesem nazývaným jako glukoneogeneze. Velkou část energie je nutné po zatížení investovat také do odstranění nadbytečného tepla (termoregulace – energeticky velmi náročný proces). Na základě pozátěžového poklesu SF (vagové reaktivace) při stejném absolutním zatížení, je možné posoudit trénovanost dvou osob, přičemž platí, že čím je rychlejší pokles SF, tím je člověk trénovanější.
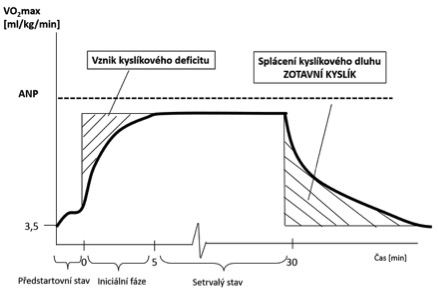
Obrázek 1.7 Vznik kyslíkového deficitu a zotavný kyslík
Svalový systém
Všechny aktivní pohyby, které člověk vykovává, jako například chůze, běh, plavání nebo jen jemné pohyby prstů či změna výrazu v obličeji, jsou výsledkem koordinované součinnosti kosterních svalů. Centrum řízení volní motoriky je lokalizováno v šedé kůře mozkové, konkrétně v motorickém centru umístěného v gyrusprecentralis(frontální lalok). Svalová kontrakce je proces, který probíhá mezi kontraktilními bílkovinami – aktinem a myozinem za dodávky energii (ATP) a přítomnosti iontů vápníku (Ca2+), které jsou skladovány v T-tubulech. Svalová kontrakce je proces, který probíhá mezi kontraktilními bílkovinami – aktinem a myozinem za dodávky energie (ATP) a přítomnosti iontů vápníku (Ca2+), které jsou skladovány v T-tubulech. Síla kontrakce se odvíjí nejen od typu aktivovaného svalového vlákna, ale také od časového průběhu zapojení motorických jednotek.
Svalová tkáň, především červená vlákna, jsou bohatě protkána systémem kapilár, které svalovým buňkám zajišťují dodávku kyslíku a živin (glukózy + VMK) a odvod z plodin metabolismu včetně CO2. V klidu je většina (95 %) z těchto kapilár zúžená až uzavřená. Zatímco při tělesné práci vlivem působení katecholaminů u nich dochází k vazodilataci a tím se zvyšuje lokální mikrocirkulace, což můžeme na svalech pozorovat po rozcvičení, kdy navýší svůj objem. Vlákno příčně pruhovaného svalu představuje obrovské soubuní délky až 15 cm, které obsahuje mnoho jader a další standardní buněčné organely. Na rozdíl od jiných buněk v organismu se však zralé svalové vlákno nemůže dále dělit, čili počet svalových vláken zůstává během života stejný. Poškozené svalové vlákno je možné nahradit díky specializovaným tzv. satelitním buňkám, aniž by došlo ke změně jeho typologie.
Kosterní (příčně pruhované) svalstvo tvoří u žen zhruba 25–35 % a u mužů 40–45 % tělesné hmotnosti. Svalová vlákna obsahují 75 % vody, 20 % bílkovin a pouhých 5 % připadá na sacharidy, tuky a anorganické soli. Základní fyziologickou vlastností svalu je stažlivost a dráždivost. Svalová vlákna v lidském organismu rozdělujeme na dva, respektive tři typy, které se od sebe navzájem liší svojí morfologií a funkčními vlastnostmi.
Prvním typem svalových vláken (typ I.) jsou pomalá nebo také červená (oxidativní) vlákna; druhým typem vláken (II. B) jsou rychlá nebo také bílá (glykolytická) vlákna a třetím typem (II. A) jsou vlákna taktéž rychlá, ale oxidativní. Ta představují jistý metabolicko-funkční mezistupeň mezi červenými a bílými vlákny. Za hlavní rozdíl mezi vlákny bývá uváděna rychlost cyklu kontrakce-relaxace. Pomalé vlákno potřebuje na dosažení maximálního zkrácení přibližně dvojnásobek času v porovnání s rychlým vláknem. Rychlost a síla svalové kontrakce je dána především typem inervace a enzymatickou výbavou vlákna. Pomalá svalová vlákna jsou inervována motoneurony s pomalým vedením vzruchu, zatímco rychlá vlákna jsou spojena s motoneurony s rychlým vedením vzruchu. Motoneurony společně se svalovými vlákny vytvářejí motorickou jednotku, kterou lze podle počtu (10 až 1000) inervovaných svalových vláken jedním motoneuronem, dělit na velkou (velká síla + hrubé pohyby, např. gluteální svaly) a malou (jemná – přesné pohyby, např. okohybné svaly) motorickou jednotku. Vlákna I. typu – červená vlákna mají v průměru 45–55% zastoupení ve svalech dolních a horních končetin. Červená vlákna mají vyšší podíl ve svalech posturálních než fázických.
Mezi základní charakteristiky pomalých vláken (typ I.) patří:
- nižší aktivita myozinové ATPázy (enzym urychlující zisk energie – štěpením ATP),
- nižší schopnost rychle přemisťovat ionty vápníku (ovlivňuje rychlost kontrakce),
- vyšší počet velkých mitochondrií,
- vyšší aktivita aerobních (mitochondriálních) enzymů,
- vysoká denzita kapilár (vyšší extrakce O2),
- vyšší rezistence vůči únavě,
- vyšší koncentrace myoglobinu,
- nižší produkce energie za jednotku času (pomalejší aerobní tvorba ATP),
- nižší síla kontrakce,
- nižší práh dráždivosti,
- nižší příčný průřez vlákna.
Pro bílá svalová vlákna (typ II. B) je typické:
- vyšší aktivita myozinové ATP-ázy,
- vyšší schopnost rychle přemisťovat ionty vápníku,
- vyšší enzymatické zastoupení glykolytických enzymů (tvorba La + ↑H+),
- zvýšený význam CP (rychlá resyntéza ATP),
- malý počet mitochondrií,
- nízké zastoupení a aktivita aerobních enzymů,
- nízká denzita kapilár (nižší extrakce O2),
- nižší rezistence vůči únavě (důsledek anaerobní produkce ATP),
- nízká koncentrace myoglobinu,
- vysoká produkce energie za jednotku času (ATP-CP + anaerobní produkce ATP),
- vyšší síla kontrakce,
- vyšší práh dráždivosti (vyšší stimulační podnět),
- vyšší příčný průřez vlákna (zvýšená tendence k hypertrofii).
Základní charakteristika svalových vláken typu II. A (rychlá – oxidativní), která jsou na pomezí funkčně i morfologicky velmi vyhraněných svalových vláken (I. a II. B), se vyznačuje tím, že některá vlákna typu II. B (rychlá – glykolytická) obsahují větší množství mitochondrií a enzymů aerobního metabolismu.
U svalové kontrakce se rozlišuje několik základních typů, které souvisí se změnou délky svalového vlákna a jeho napětí (tonu) při provádění pohybu.
Prvním typem je izometrická svalové kontrakce, při které se aktivovaný sval snaží zkrátit, ale zkrácení je bráněno například fixací nebo stejně silnou snahou o jeho protažení. Ve výsledku dochází ke zvýšení svalového tonu, který však není doprovázen změnou délky svalu. Druhý typ svalové kontrakce reprezentuje izotonická kontrakce, která je podle literatury spíše teoretickou situací, protože by znamenala zkrácení svalu při neměnném svalovém napětí. Dále rozeznáváme koncentrickou a excentrickou svalovou kontrakci. Při koncentrické kontrakci dochází ke zkracování délky svalu a překonávání gravitace či odporu břemene. Naopak u excentrické kontrakce nastává postupné protahování svalu, protože externí síla působící na sval je větší než síla vyvíjená při svalové kontrakci. Excentrická kontrakce je též nazývána jako brzdivá (decelerační). Během protahování svalového vlákna včetně šlach a vazů dochází v těchto strukturách k postupné akumulaci elastické energie. Tuto energii je možné následně využít pro zvýšení celkové intenzity svalového stahu. Tento princip funguje pouze za předpokladu, že po excentrické kontrakci dojde k okamžitému navázání aktivní koncentrickou kontrakcí bez poklesu svalového napětí.
Kontrolní otázky
- Čím je zprostředkována regulace dýchání?
- Na základě jakého principu dochází k výměně plynů mezi plicemi a krví?
- Které faktory ovlivňují disociační křivku kyslíku?
- Co je to minutový srdeční výdej?
- Jak lze stanovit maximální srdeční frekvenci?
- Co je to arteriovenózní diference kyslíku?
- Jak lze charakterizovat setrvalý stav?
- Co je to kyslíkový deficit?
- Jaké jsou charakteristické vlastnosti svalového vlákna typu II. B?
- Jaký je rozdíl mezi excentrickou a koncentrickou svalovou kontrakcí?
Literatura
Åstrand, P.-O., Rodahl, K., Dahl, H., & Strømme, S. B. (2003). Textbook of work physiology: Physiological bases of exercises (4th ed.). Champaign, IL: Human Kinetics.
Brooks, G. A., Fahey, T. D., & White, T. P. Baldwin, K. M. (2000). Exercise physiology: Human bioenergetics and its applications (3rd ed.). New York, NY: McGraw-Hill Companies.
Ganong, W. F., Herget, J., Blažek, T., & Herget, J. (1995). Přehled lékařské fyziologie. Jinočany: H&H.
Hamar, D., & Lipková, J. (2001). Fyziológia telesných cvičení. Bratislava: Univerzita Komenského.
Máček, M., & Máčková, J. (1995). Fyziologie tělesných cvičení. Praha: SRZTV.
Máček, M., & Radvanský, J. (2011). Fyziologie a klinické aspekty pohybové aktivity. Praha: Galén.
Máček, M., & Vávra, J. (1988). Fyziologie a patofyziologie tělesné zátěže. Praha: Avicenum.
Rokyta, R. et al. (2000). Fyziologie. Praha: ISV.
Trojan, S. (2003). Lékařská fyziologie. Praha: Grada Publishing.