Fyziologické determinanty vytrvalostního výkonu
Vytrvalost je obecně chápána jako schopnost dlouhodobě provádět tělesnou práci určité intenzity bez snížení její efektivity. Vytrvalost lze dělit podle doby trvání na rychlostní ( sprinterskou) (7–35 s); krátkodobou (35 s až 2 min), střednědobou (2–10 min) a dlouhodobou (> 10 min). Z hlediska metabolického krytí se anaerobní metabolismus do resyntézy ATP bude nejvíce zapojovat při rychlostní a do 60 s výkonu také při krátkodobé vytrvalosti, nicméně u střednědobé a dlouhodobé vytrvalosti už ATP pochází téměř výlučně z aerobních procesů.
Mezi významné determinanty dosažení vysoké úrovně vytrvalostního výkonu patří:
1. typologie svalových vláken,
2. VO2max,
2A. úroveň AP a ANP
3. ekonomika pohybu.
Typologie svalových vláken
Procentuální zastoupení jednotlivých typů příčně pruhovaných svalových
vláken je individuálně variabilní a na této variabilitě má největší podíl
genetika. Uvádí se, že v populaci se nejčastěji vyskytuje poměr 45 % pomalých
a 55 % rychlých svalových vláken. Nicméně platí, že počet jedinců s dominantním
zastoupením pomalých nebo rychlých svalových vláken je tím menší, čím více
se blíží k jednomu či druhému extrému. Zjišťování podílu jednotlivých typů
svalových vláken a tedy i jisté předpoklady, zda sportovec bude výkonnostně
inklinovat spíše k rychlostně-silovým nebo vytrvalostním disciplínám, se
dříve v hojném počtu (aktuálně spíše v nižším počtu a zejména pro vědecké
účely) provádělo pomocí
svalové biopsie
(odběr vlákna např. z vastus lateralis). Vzhledem k tomu, že svalová
vlákna jsou svojí výbavou funkčně natolik specializovaná, je zcela logické,
že vytrvalci budou disponovat dominantně oxidativními vlákny. V důsledku
specifických adaptačních změn vyplývajících z délky a intenzity tréninkového a
závodního zatížení, je převaha oxidativních vláken nad rychlými glykolytickými
ještě umocňována podílem vláken typu II. A, tedy rychlými oxidativními.
U elitních vytrvalců se uvádí až 80% zastoupení pomalých oxidativních vláken oproti 20% zastoupení rychlých glykolytických.
Dominantní podíl oxidativních svalových vláken, která patří k největším
konzumentům kyslíku během tělesné práce, jenom zvýrazňuje velké možnosti
aerobního zisku energie (ATP) kterým vyspělí vytrvalci disponují. Vlivem
vysoké oxidační kapacity svalových vláken mohou vytrvalostně trénovaní
jedinci ve větší míře využívat energii pro svalovou činnost pocházející
ze štěpení
tuků, respektive volných mastných kyselin. Pro svalová vlákna typu I. je dále typické, že laktát vlivem vysoké
oxidativní kapacity vytváří pouze omezeně a spíše se podílí na jeho zpracovávání
(metabolizaci).
Maximální spotřeba kyslíku
Maximální spotřeba kyslíku je definována jako maximální množství z přijatého kyslíku, který je organismus schopen využít při maximální svalové práci. Bývá označována zkratkou VO2max a můžeme ji vyjádřit v absolutních (L/min) nebo v relativníchjednotkách(ml/kg/min).
Uvádí se, že pracovat na intenzitě odpovídající úrovni VO2max lze pouze po relativně omezenou dobu, která se u vyspělých vytrvalců pohybuje kolem 10 až 15 minut. Intenzita zatížení (vyjádřeno v procentech VO2max) negativně koreluje s délkou jejího trvání. Svou roli zde sehrává i trénovanost jedince, přičemž platí, že trénovanější osoba je schopna s rostoucí dobou trvání tělesné práce udržovat vyšší intenzitu zatížení ve srovnání s osobou méně trénovanou. Bylo zjištěno, že u nedostatečně trénované osoby se podíl využití VO2max při různé době trvání zatížení oproti trénovanému jedinci snižuje o 20 až 30 %. U dvou vytrvalců se shodnou hodnotou VO2max pak o maximální udržitelné rychlosti běhu rozhoduje úroveň ANP.
Pro zjišťování vytrvalostních předpokladů se častěji využívá relativního vyjádření VO2max, tedy s přepočtem na kg hmotnosti, které umožňuje vzájemné porovnávání sportovců s odlišnou tělesnou hmotností. Vyšší VO2max ukazuje na lepší dispozice pro intenzivnější vytrvalostní zatížení a tedy i pro vytrvalostní výkon, zatímco patologicky nízké hodnoty VO2max mohou začít člověka limitovat i při relativně nenáročných pohybových aktivitách habituálního charakteru. Platí, že čím vyšší bude množství kyslíku, které je organismus schopen během svalové práce využít, tím více energie bude vytvářeno efektivním aerobním způsobem. Ukazatel VO2max má největší diagnostický význam zejména za podmínek zapojení co největšího množství svalových skupin během dynamické práce. Uvádí se, že hodnoty VO2max zjišťované při spiroergometrii na běhátku jsou v průměru o 5–10 % (výjimečně až o 20 %) vyšší vzhledem k hodnotám VO2max zjištěné na bicyklovém ergometru. Nicméně, u vysoce vytrvalostně trénovaných jedinců, jako jsou například běžci na střední či dlouhé distance, silniční cyklisté,–nebo chodci, budou naměřeny vyšší hodnoty VO2max, když tito sportovci budou testováni při jejich specifickém pohybu čili např. cyklista na bicyklovém ergometru nebo veslař na veslařském trenažéru. Zvýšené hodnoty VO2max při testování za specifických pohybových podmínek mohou být výsledkem tzv. lokální trénovanosti. Stanovení VO2max má však pouze prediktivní charakter s ohledem na vytrvalostní výkon, protože reálná vytrvalostní výkonnost v sobě více zohledňuje ekonomiku pohybu a úroveň ANP. Kromě indikace vytrvalostních schopností lze považovat hodnotu VO2max za ukazatel zotavných procesů při intermitentních pohybových aktivitách (např. sportovních hrách), přičemž se má za to, že sportovec s vyšší aerobní kapacitou je schopen absolvovat více sprintů a lépe odolávat nástupu únavy než sportovec s nižší VO2max díky rychlejšímu splácení kyslíkového dluhu vytvořeného během vysoce intenzivních fází zatížení. U mladých netrénovaných mužů, resp. žen se udávají hodnoty VO2max okolo 3 L/min, resp. 2 L/min, které při vztažení na kilogram hmotnosti (muž 70 kg, žena 55 kg) činí u mužů okolo 45 ml/min/kg a 36 ml/min/kg u žen. Za vynikající hodnoty VO2max u mužů resp. u žen lze považovat VO2max > 70 ml/min/kg; resp. > 60 ml/min/kg. U extrémně vytrvalostně trénovaných jedinců nejsou výjimkou hodnoty 5 až 7 L/min, což v závislosti na hmotnosti sportovce mohou být hodnoty mezi 80 až 90 ml/min/kg. Tak vysoké hodnoty VO2max jsou mezi sportovci dosahovány většinou běžci na lyžích, kteří kromě toho, že při pohybu zapojují velký počet svalových skupin, absolvují i mnoho tréninků ve vyšší nadmořské výšce, což přispívá k vysoké aerobní kapacitě.
Výslednou hodnotu VO2max ovlivňuje více fyziologických parametrů. Na začátku celého řetězce stojí dýchací systém, kde na úrovni plicních sklípků, resp. alveokapilární membrány, dochází na základě rozdílného parciálního tlaku O2 k difuzi kyslíku do krve, která kyslík transportuje k pracujícím svalům. Jakmile se krev dostane do kapilární sítě svalu, část kyslíku se zde uvolní z vazby na hemoglobin a je využita enzymy aerobního metabolismu v mitochondrii (v dýchacím řetězci) pro oxidaci vodíku za vzniku tzv. metabolické vody. Ke snadnějšímu uvolnění vazby kyslíku z hemoglobinu napomáhá na úrovni svalového vlákna při intenzivní práci posun disociační křivky kyslíku směrem doprava vlivem zvýšené teploty (vznik při kontrakci), poklesem pH (vzestupem H+), zvýšeným pCO2 (tzv. Bohrův efekt) a zvýšenou koncentrací 2,3 DPG. Jako předpoklad vysoké aerobní výkonnosti se uvádí efektivní a účinná regulace svalové mikrocirkulace. Zbytek nevyužitého kyslíku se společně s vytvořeným CO2 vrací žilním systémem zpět do srdce a malým plicním oběhem do plic, kde je CO2 vydýchán. Jak již bylo podrobněji vysvětleno v kapitole věnované transportnímu systému, rozdíl mezi obsahem kyslíku v arteriální a žilní krvi se nazývá arteriovenózní diference kyslíku (a-v)O2. Za limitující faktor VO2max se u zdravého jedince považuje výkonnost srdce a to ve smyslu omezených možností přečerpaného objemu krve společně se schopností extrahovat kyslík z protékající krve kapilárním systémem svalu. Společně se zvýšeným objemem cirkulující krve a zvýšenou výkonností srdce je právě zvýšená extrakce kyslíku považována za významný projev adaptace při vytrvalostním tréninku přispívající k vzestupu hodnoty VO2max. V posledních letech bylo u extrémně vytrvalostně trénovaných sportovců (VO2max > 75 ml/min/kg) zjištěno, že limitujícím faktorem může být i doba alveokapilární difuze kyslíku, protože za podmínek vysoké výkonnosti srdce (vysoký MSV) už krev proudí plicemi natolik rychle, že se nestačí plně saturovat kyslíkem. Spotřebu kyslíku lze stanovit pomocí Fickovy rovnice:
VO2 = Q (SF · SV) · (a-v)O2,
kde Q – minutový srdeční výdej (L/min), SF – srdeční frekvence (tep/min), SV –systolický objem (ml), (a-v)O2 – arteriovenózní diference kyslíku (ml).
Výsledná hodnota VO2max se odvíjí od velikosti orgánů participujících na transportu kyslíku (plíce, srdce, krev) a typologii svalových vláken. Z tohoto pohledu budou dosahovat vyšších hodnot VO2max jedinci robustnější než gracilnější, ale při přepočtu VO2max na kg hmotnosti se rozdíly stírají nebo dokonce mohou mít gracilnější jedinci VO2max i vyšší. Vysoká hodnota VO2max (kg/min) není u dobře trénovaných vytrvalců pouze výsledkem dlouhodobého tréninku, jak je mnohdy prezentováno, protože prostor pro zvyšování VO2max tréninkem je relativně hodně omezený. V současnosti je v souvislosti s odezvou VO2max na aerobní trénink zmiňována vysoká interindividuální variabilita ve změně aerobní kapacity, která se pohybuje v rozsahu od 0 do 40 %, ale průměrně se udává zvýšení okolo 25 % u netrénované populace. Vysoká variabilita v odezvě VO2max na trénink je přisuzována genetickým dispozicím. Příklad dobrých genetických předpokladů pro vytrvalostní výkon reprezentují Keňané, kteří již po celé generaci žijí ve vyšší nadmořské výšce. U těchto jedinců lze vytrvalostním tréninkem dále zvyšovat již přirozeně vysokou hodnotu VO2max a vytvářet tak funkční předpoklady pro vysokou vytrvalostní výkonnost. Naproti tomu u jedince s nízkým genetickým tréninkovým potenciálem a podprůměrnou vstupní hodnotou VO2max nelze ani při sebeintenzivnějším tréninku očekávat markantní zvýšení VO2max. V poslední době se ukazuje, že geneticky není podmíněna pouze vstupní úroveň VO2max, ale i velikost jejího přírustku související s trénovatelností (adaptabilitou) jedince. Termín trénovatelnost (adaptabilitu) je možné charakterizovat jako geneticky podmíněnou schopnost organismu přizpůsobit se stresovým (tréninkovým) podnětů, které vytvářejí základ pro vznik morfo-funkčních adaptací. Adaptabilita bývá často asociována s regulační kapacitou ANS, konkrétně s aktivitou parasympatiku (n. vagu). V kontextu zvyšování vytrvalostních schopností a aktivity vagu, lze konstatovat, že sportovci s vyšší vagovou regulací mají výhodnější podmínky pro zvyšování hodnoty VO2max (ml/kg/min), než sportovci s přirozeně nižší kardiální vagovou regulací.
Rozhodující podíl zlepšení VO2max v průběhu tréninku je identifikovatelný už v prvních měsících tréninku. V pozdějších fázích se hodnota VO2max stabilizuje, ale někdy i mírně klesá, proto je podle některých autorů hodnocení efektivity tréninkového procesu především u vytrvalců pouze na základě změny v hodnotě VO2max přinejmenším ne zcela objektivní.
Při zjišťování hodnoty VO2max, by se měla testovaná osoba postupným zvyšováním zatížení vystavit podmínkám úplného vyčerpání, které je však do jisté míry záležitostí subjektivního posouzení každého testovaného jedince. Z tohoto důvodu byly ustanoveny všeobecně platná kritéria pro objektivizaci dosažení podmínek vita maxima, kterými jsou:
- Maximální srdeční frekvence (SFmax) – při ukončení práce by měla SF dosahovat aspoň 85 % hodnoty z věkově predikovaného maxima, čili SFmax ≥ (220 – věk) · 0.85.
- Respirační kvocient (RQ) – hodnota RQ by měla být vyšší než 1,10, někdy se uvádějí i hodnoty 1,14 až 1,17. Vzestup RQ nad 1 (hodnota RQ sacharidů) je od určité intenzity zatížení výsledkem významného zapojení anaerobního metabolismu do krytí energie (ATP) pro pracující svaly, přičemž vzniká H+ (pokles pH), který je působením pufrovacího systému neutralizován za vzniku CO2 a H2O. CO2 je ve zvýšené míře v plicích vydýcháván (hyperventilace), což se projeví ve vzestupu RQ. Hodnoty RQ>1,10 nemusí být striktně dosaženo u velmi dobře vytrvalostně trénovaných jedinců s vysokou hodnotou VO2max z důvodu nižší anaerobní kapacity (nízká produkce laktátu a H+).
- Vznik plató (leveling off) u VO2 – v poslední fázi dynamické práce by mělo nastat tzv. plató ve spotřebě kyslíku, tedy hodnota VO2 by ještě i přes krátkodobé pokračování v zátěži měla stagnovat, popřípadě i mírně klesat.
- Hladina laktátu – po zatížení by měla koncentrace laktátu přesahovat 7 až 10 mmol/L.
- Ventilační ekvivalent pro kyslík (V/VO2) – hodnota na konci zatížení by měla přesahovat 35 L.
Aerobní práh
Při zvyšující se intenzitě zatížení dochází k zvyšování nároků na dodávku energie pro pracující svaly. Čím více roste intenzita zatížení, tím více se do pohybů začínají zapojovat rychlá svalová vlákna a roste produkce laktátu a H+. Aerobní práh (AP) je prvním ze dvou metabolických předělů, přičemž jedna z definic o AP hovoří jako o intenzitě zatížení, kdy se energie pro svalovou činnost (ATP) přestává tvořit výlučně pomocí aerobních procesů, což se projeví narušením normální hladiny laktátu v krvi. Za hladinu laktátu, která ukazuje při zvyšujícím se zatížení na úroveň AP, se obecně považuje koncentrace do 2 mmol/L (obrázek 1.11). U běžné netrénované populace ve věku od 18 do 30 let odpovídá hranici AP intenzita zatížení zhruba 50 % VO2max. Úroveň AP se při tréninku zejména kolektivních sportů využívá víceméně sporadicky, slouží především pro účely regeneračních tréninků. Významnější roli hraje úroveň AP u individuálních sportů vytrvalostního charakteru jako je například silniční cyklistika, triatlon nebo běhy na dlouhé distance, ve kterých se velká část objemových tréninků odehrává právě na hranici AP. U cyklistů je udávána hranice AP někde kolem 60–65 % VO2max, kde se zároveň nachází u vytrvalostně trénovaných jedinců hranice nejvyššího mitochondriálního obratu VMK.
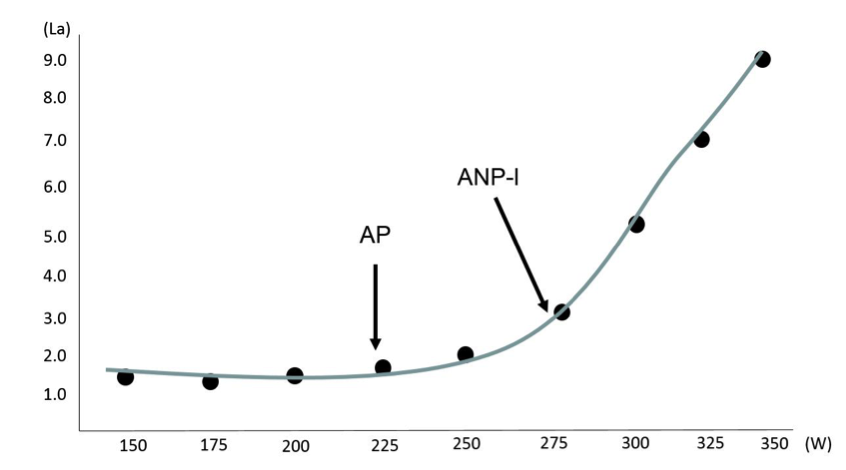
Obrázek 1.11 Konstrukce laktátové křivky a determinace aerobního a anaerobního prahu
Anaerobní práh
Schopnost využít během déletrvajícího vytrvalostního výkonu co nejvyšší podíl VO2max těsně souvisí s úrovní ANP. Determinace ANP má pro zvyšování výkonnosti u většiny sportů větší význam než znalost hranice AP. V literatuře se rozeznávají tři druhy ANP:
- ANP cirkulační (srdeční frekvence) – neinvazivní stanovení ANP
- ANP metabolický (koncentrace laktátu) – invazivní stanovení ANP
- ANP ventilační (dechové plyny) – neinvazivní stanovení ANP
Ad1) ANP cirkulační (ANP-c) se stanovuje na základě stupňovaného testu do maxima. Často používaným testem pro determinaci ANP-C je protokol Conconiho testu, který je znám v několika modifikacích podle druhu sportu (např. pro běh, cyklistiku nebo plavání). Principem testu je sledování odezvy SF na zvyšující se zatížení, přičemž mezi zatížením a SF od 110–120 tep/min existuje lineární vztah. Od určitého momentu je možné při kontinuálním zvyšování zatížení identifikovat odklon SF od doposud udržované linearity, který se nazývá ANP-C (obrázek 1.12). Od tohoto momentu se srdeční frekvence zvyšuje při rostoucím zatížení již pozvolněji. Uvádí se, že ANP-C se nachází u osob mezi 18–30 rokem věku i přes inter individuální variabilitu v rozmezí 170–180 tep/min. Bylo zjištěno, že zlom SF na úrovni ANP-C je snadněji identifikovatelný u vytrvalostně trénovaných sportovců než u běžné populace.
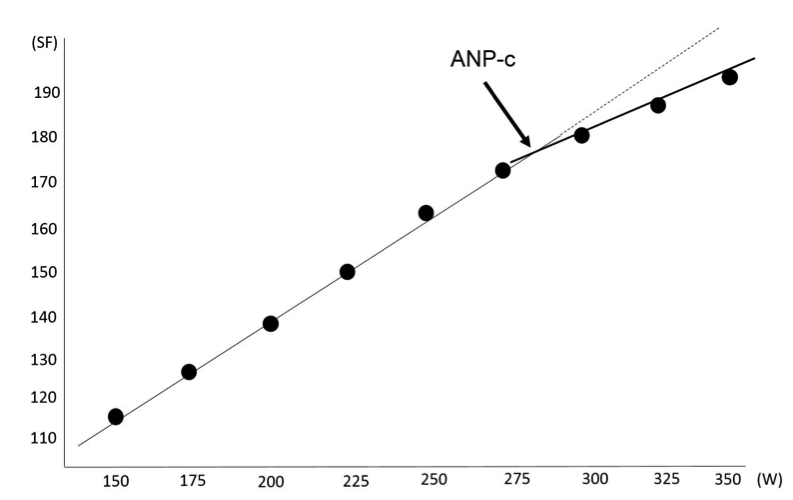
Obrázek 1.12 Ukázka determinace cirkulačního anaerobního prahu
Ad2) ANP metabolický neboli laktátový ANP (ANP-l) se definuje jako intenzita zatížení (vyjádřená např. v % VO2max nebo rychlostí běhu), při které dochází k narušení dynamické rovnováhy mezi produkcí laktátu a schopností jeho odplavení nebo využití jako zdroje pro svalovou činnost. Výsledkem narušení rovnováhy je kumulace laktátu a vodíkových iontů (H+), které jsou zodpovědné za pokles pH a zvyšující se únavu. ANP-l se zpravidla stanovuje na základě laktátové křivky, která se konstruuje pomocí několika stupňového zatížení s rostoucí intenzitou. Každý stupeň trvá nejméně 4 minuty, přičemž jsou známy modifikace, kde jeden stupeň trvá i dvojnásobný čas. Na konci každého stupně, kterých podle druhu protokolu bývá od 3 do 8, se z bříška prstu nebo ušního lalůčku odebírá vzorek kapilární krve pro analýzu koncentrace laktátu. Hladina laktátu se v důsledku zvětšujícího se podílu anaerobní tvorby energie během stupňujícího se zatížení zvyšuje z klidové úrovně 0,80–1,50 mmol/L nejprve pozvolně, protože vyprodukovaný laktát je tělo schopno zpracovat jako energetický substrát. V okamžiku, kdy je porušena dynamická rovnováha mezi produkcí a zpracováním laktátu se jeho koncentrace začne zvyšovat exponenciálně. Tato intenzita zatížení odpovídá ANP-L. Podle aktuální literatury se hodnota ANP-L nachází od 2.5 do 6.0 mmol/L. Při konstrukci laktátové křivky bude možné pozorovat rozdíly v kinetice laktátu podle sportu, který sportovec vykonává. U sprintera či čtvrtkaře se hladina laktátu bude zvyšovat již při nižších intenzitách, zatímco u vytrvalostně trénovaných sportovců bude vzestup laktátu pozorovatelný až při vyšším zatížení. Dříve se hodnota ANP stanovovala při koncentraci laktátu v žilní či kapilární krvi na hranici 4 mmol/L, což znamenalo metabolické rozhraní mezi aerobním a výhradně anaerobním vznikem ATP. Na základě nových poznatků a sofistikovanějších metod hodnocení ANP se postupně upustilo od obecně udávané hodnoty 4 mmol/L a začal se stanovovat tzv. individuální ANP (ANPi), který se nachází v rozmezí 3 až 6, eventuálně až 8 mmol/L. Se značnou variabilitou hladiny laktátu na ANP souvisí relativně nový pojem zavedený do metodiky hodnocení laktátu, kterým je maximální laktátový setrvalý stav (MaximalLactatSteadyState – MLSS). Jedná se o nejvyšší možnou koncentrace laktátu udržitelnou během dynamické svalové práce(balance mezi produkcí a utilizací laktátu aerobními procesy), která nastává při intenzivní práci (50–80 % VO2max) zhruba po 10 až 15 min zatížení. Bylo prokázáno, že hodnota laktátu na úrovni MLSS nemá žádnou souvislost se zdatností nebo sportovní výkonností. Při déletrvajícím vytrvalostním zatížení tedy není primárně rozhodující úroveň hladiny laktátu na ANP, která u dobře aerobně disponovaných sportovců může být odlišná, ale podávaný výkon (W) nebo rychlost běhu na úrovni ANPi, resp. MLSS, který se spíše odvíjí od schopnosti udržet stabilní nitrosvalové pH (H+) než od hladiny laktátu samotné. Z výše uvedeného je zřejmé, že intenzita zatížení odpovídající ANP-l je mimo jiné ovlivňována faktory, jako je například:
- podíl červených (oxidativních) vláken – významný konzument laktátu,
- hustotou kapilární sítě ve svalu – vyšší hustota zvyšuje dostupnost kyslíku pro aerobní pochody,
- aktivitou aerobních enzymů,
- počtem a hustotou mitochondrií,
- schopností pufrovacího systému eliminovat H+.
Úroveň ANP-l se u netrénované populace nachází okolo
60 až 70 % VO2max, zatímco vlivem tréninku se u vysoce vytrvalostně trénovaných sportovců
mohou hodnoty ANP blížit hranici
90 % VO2max. Výhodou hodnocení úrovně ANP během tréninkového procesu je
její větší dynamika než u hodnoty VO2max. Pozornost se však upírá nikoliv na hladinu laktátu, ale jak již bylo
zmíněno výše, na výkon nebo na rychlost běhu popřípadě na úroveň %VO2max odpovídající ANP. V případě dvou vytrvalců s téměř shodnou hodnotou VO2max
= 81 a 79 ml/kg/min, ale odlišnou úrovní ANP – 78 a 89 %VO2max
(63 a 70 ml/kg/min), je možné pro lepší vytrvalostní výkon spíše favorizovat
druhého sportovce s výše položeným ANP.
ad3) ANP ventilační (ANP-v) se určuje na základě hodnocení změn v dynamice výměny dechových plynů (kyslíku a oxidu uhličitého) při stupňovaném zátěžovém protokolu. Ventilační odpověď organismu při stupňovaném zatížení souvisí s rostoucím podílem anaerobního metabolismu na hrazení energie pro pracující svaly. Vlivem zvyšující se produkce laktátu a také iontů H+, které jsou zodpovědné za pokles pH, dochází k úpravě porušené acidobazické rovnováhy pomocí pufrovacího systému uhličitanové, fosforečnanové nebo bílkovinné povahy. Navázáním volného H+ na uhličitanový pufr dochází ke zvyšování parciálního tlaku CO2 v krvi (obrázek 1.13), který následně stimuluje dýchací centrum uložené v prodloužené míše a dochází k intenzivnější ventilační odpovědi (hyperventilaci) s cílem redukovat nadbytečné množství CO2 v organismu a stabilizovat pH. Úroveň ANP-v se obvykle určuje z grafu, kde je vykreslena dynamika minutové ventilace, spotřeby kyslíku, výdeje oxidu uhličitého. ANP-v pak odpovídá bodu, kdy dochází k náhlému zvyšování výdeje oxidu uhličitého a odchylování ventilace od paralelního vzestupu spotřeby kyslíku jako odpověď na metabolické změny související s kumulací H+ a činností pufrů.
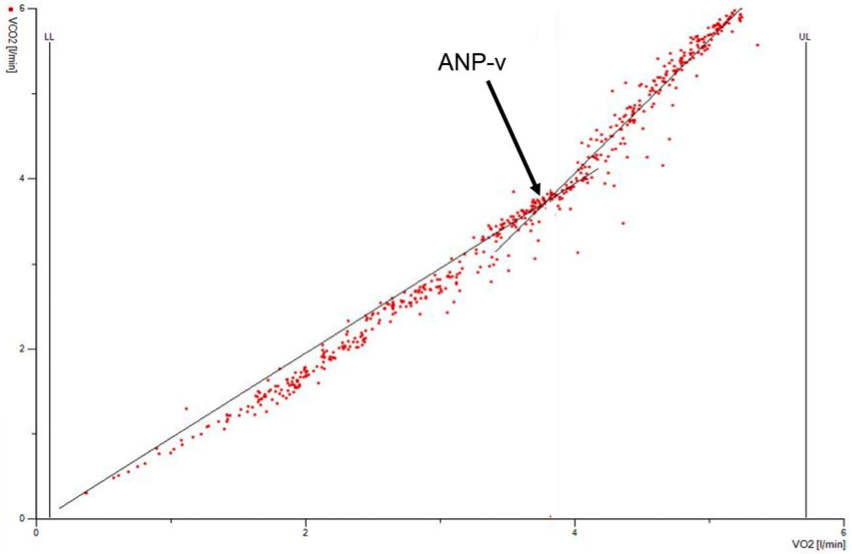
Obrázek 1.13 Ukázka determinace ventilačního anaerobního prahu
Využití anaerobního prahu v tréninku
Vlivem vytrvalostního tréninku dochází ke zvyšování aerobní kapacity a tím i úrovně ANP. U vyspělých vytrvalců však v důsledku tréninku nedochází k tak významným změnám v hodnotě VO2max, ale za indikátor změny úrovně trénovanosti slouží spíše úroveň ANP, respektive výkon podávaný na ANP. Vlivem tréninku dochází k posunu ANP-L směrem doprava, což například u cyklisty znamená, že došlo ke zvýšení výkonu vyjádřených ve wattech na ANP (obrázek 1.14). Jednou z možností, jak přistupovat k rozvoji aerobní kapacity a tím i ke zvyšování úrovně ANP, je trénink na intenzitě těsně pod úrovní ANP, přičemž praxe ukazuje, že takto vysokou intenzitu zatížení jsou schopni vytrvalci tolerovat přibližně 40–60 min a navíc za cenu velkého úbytku svalového glykogenu. Obnova glykogenových zásob se odhaduje zhruba na 48 hodin, proto se tento typ tréninku doporučuje aplikovat maximálně 2–3x v týdnu, aby nedocházelo k poklesu kvality tréninku z důvodu nedostatku svalového glykogenu. Pro rozvoj aerobní, ale převážně anaerobní kapacity je typické spíše než kontinuální zatížení, použít zatížení intervalové, při kterém se intenzita zatížení opakovaně nachází nad úrovní ANP a dochází tak k opakované stimulaci anaerobního metabolismu.
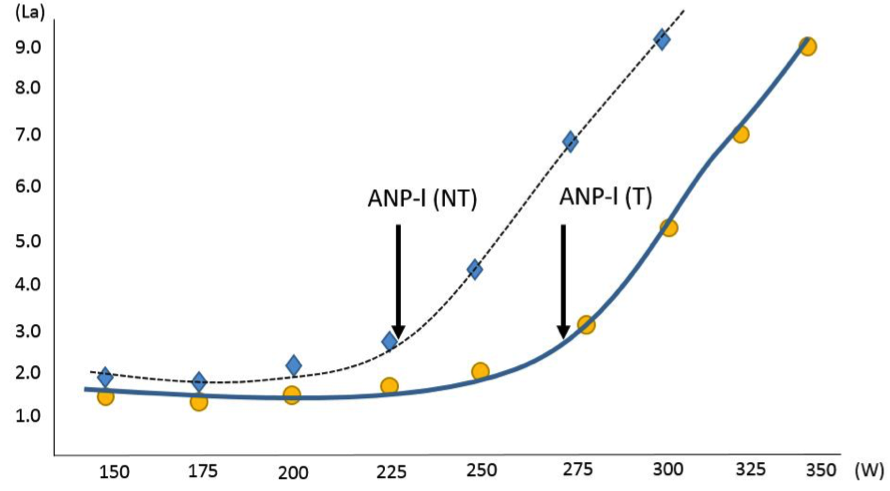
Obrázek 1.14 Efekt vytrvalostního tréninku na úroveň anaerobního prahu
Ekonomika pohybu
Ekonomika pohybu bývá často vyjadřována pomocí množství spotřebovaného kyslíku (ml/kg/min) při dané rychlosti pohybu na ergometru. Při zvyšujícím se zatížení roste lineárně VO2, ale u sportovce s horší ekonomikou pohybu bude při stejné rychlosti spotřebováno větší množství energie (vyšší hodnota VO2), proto teoreticky při dosažení shodné VO2max poběží sportovec s horší ekonomikou pohybu pomaleji než sportovec s lepší ekonomikou pohybu. Ekonomika pohybu je považována za multifaktoriální veličinu. Za jeden z hlavních faktorů ovlivňující ekonomiku pohybu jsou považovány tělesné parametry běžce, přičemž se za ideální považuje velká tělesná štíhlost, zejména dolních končetin (lýtek), a nižší vzrůst (etiopští nebo keňští běžci). Ekonomika pohybu je také významně ovlivněna technikou běhu, přičemž mezi technikou a ekonomikou platí positivní vztah. V neposlední řadě je nutno uvést také endogenní faktory, jako např. odpor vzduchu, který ekonomiku pohybu může zásadním způsobem ovlivnit (např. běh nebo jízda na kole v tzv. vzduchovém pytli).
Fyziologické determinanty rychlostního a silového výkonu
Rychlost
Rychlost je ze všech kondičních schopností nejvíce geneticky podmíněna.
Mezi významné determinanty rychlosti patří:
- Typologie svalových vláken
- Centrální nervový systém
Typologie svalových vláken
Genetické dispozice se promítají převážně do podílu rychlých glykolytických (II. B) svalových vláken, přičemž jejich podíl se odhaduje u nejrychlejších atletů planety až na 80 %. Základní charakteristiky vláken typu II. B byly probrány v kapitole Funkční charakteristika transportního a svalového systému. Kromě typologie svalových vláken podmiňuje kvalitní sprinterský výkon i schopnost produkovat v relativně krátkém čase adekvátní množství ATP nezbytné pro svalovou práci. S ohledem na intenzitu zatížení a dobu trvání výkonů u běžců na krátké distance je produkce ATP zajišťována nejprve štěpením makroergních fosfátů (komplex ATP+CP), které však vydrží při maximální práci asi na 2 sekundy. V dalších fázích výkonu se na produkci ATP výhradním způsobem podílí anaerobní štěpení sacharidů, které katalyzují glykolytické enzymy jako je například fosfofruktokináza nebo laktátdehydrogenáza.
Centrální nervový systém
Významný podíl na rychlostním výkonu má i kvalita CNS a rychlost vedení vzruchů, které ovlivňují rychlost kontrakce a relaxace. Oproti ostatním kondičním schopnostem klade rychlost zvýšené nároky na koordinaci agonistů s antagonisticky působícími svaly, ale také na zapojování synergistů.
Síla
Sílu lze definovat jako schopnost neuromuskulárního systému vyprodukovat v daném čase co největší svalový stah. Na základě poznatků zátěžové fyziologie o zapojení typologicky odlišných svalových vláken při změně v nervové impulzaci, která je determinujícím faktorem pro konečnou úroveň vyprodukované síly, bývají podle různých autorů silové schopnosti děleny do čtyř základních druhů síly, a to na maximální sílu, rychlou (startovní a explozivní)..., reaktivní sílu a silovou vytrvalost.
Z fyziologického hlediska je zvyšování úrovně svalové síly (adaptace na silový podnět) kombinací neurálních, morfo-funkčních i metabolických změn. Morfo-funkční změny se odvíjejí od velikosti produkce anabolických hormonů, která je do jisté míry podmíněna geneticky, ale také typem silového tréninku. Za klíčové hormony pro růst svalové hmoty se považují proteosyntetické hormony: testosteron (varlata) a jeho prekurzor dehydroepiandrosteron (kůra nadledvin, v menší míře varlata a vaječníky, regulace přes adrenokortikotropní hormon – adenohypofýza), růstový hormon (adenohypofýza), inzulin (β buňky Langerhansových ostrůvků slinivky břišní), popřípadě IGF-1. Trénovatelnost síly ve smyslu jejího zvyšování je v porovnání např. se zvyšováním úrovně rychlosti relativně vysoká. Princip zvyšování objemu svalové hmoty vlivem specifického silového tréninku spočívá v poškození myofibril za vzniku mikrotraumat, která jsou během zotavení pomocí bílkovin reparována (zhojována). K výstavbě poškozených myofibril slouží specializované tzv. satelitní buňky, které se vyskytují na povrchu svalových vláken. Zvětšení objemu kontraktilních bílkovin ve svalovém vláknu se projevuje zvýšením příčného průřezu vlákna (nastává zejména u II. B) – svalovou hypertrofií.
Svaly jsou pod přímým vlivem aktivity CNS, který řídí a moduluje každou jeho funkci. Jelikož svalová kontrakce se řídí striktně podle principu "všechno nebo nic", musí být dosaženo určité minimální hraniční intenzity dráždivého impulsu proto, aby došlo ke kontrakci svalového vlákna. Další zvyšování intenzity podráždění již svalovou sílu nezvyšuje.
Organismus však disponuje mechanismy, kterými je schopen regulovat sílu svalové kontrakce (obrázek 1.15):
- zvyšování frekvence dráždivých impulsů,
- zvyšování počtu současně se kontrahujících svalových vláken.
ad 1)
Svalové vlákno reaguje na jediný akční potenciál tzv.
svalovým trhnutím. Uvádí se, že sval dosahuje zkrácení či napnutí zhruba po 80 ms po přenosu
akčního potenciálu na nervosvalovou ploténku. Jestliže dojde ke zvyšování
frekvence nervových impulsů na neuromuskulární ploténku, dochází k
nasedání další kontrakce na předchozí nedokončenou
relaxaci
a vzniká
superpozice stahů. Tento mechanismus vychází z rozdílné doby mezi kontrakcí a relaxací,
přičemž čas svalové relaxace je zhruba 10x delší než doba kontrakce. Během
superpozice stahů vzniká
vlnitý tetanus. Jestliže však nervové podněty přicházejí ještě
za vzestupné fáze kontrakce, dochází k sumaci a vzniku
hladkého tetanu. Výsledná síla během hladké tetanické kontrakce může být i více jak dvakrát
tak velká.
ad 2)
Síla svalové kontrakce se kromě časové sumace může zvýšit i
počtem aktivovaných svalových vláken tzv. prostorovou sumací.
Zvyšování nebo snižování počtu současně aktivovaných svalových vláken
není možné provádět vlákno po vlákně, protože vlákna spolu s nervovým zakončením
tvoří motorickou jednotku, tzn., že regulace aktivace či deaktivace může
probíhat pouze na
úrovni motorických jednotek.
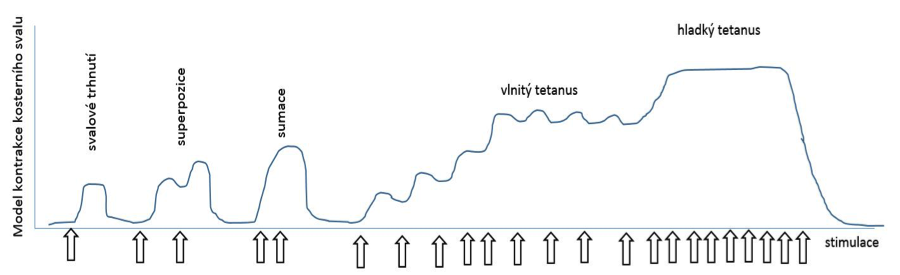
Obrázek 1.15 Regulace velikosti svalové kontrakce
Kontrolní otázky
- Jaký typ svalových vláken bude mít rozhodující podíl na vytrvalostním a rychlostním výkonu?
- Co je to maximální spotřeba kyslíku?
- Jaké druhy anaerobních prahů rozeznáváme?
- Jaké faktory ovlivňují úroveň ANP-l ?
- Které fyziologické faktory se podílejí na zvyšování úrovně síly?
- Ve kterých sportech se při řízení kondičního tréninku využívá aerobního prahu?
Literatura
Åstrand, P.-O., Rodahl, K., Dahl, H., & Strømme, S. B. (2003). Textbook of work physiology: Physiological bases of exercises (4th ed.). Champaign, IL: Human Kinetics
Beneke, R., Hütler, M., & Leithäuser, R. M. (2000). Maximal lactate-steady-state independent of performance. Medicine and Science in Sports and Exercise, 32(6), 1135–1139.
Billat, V. L., Sirvent, P., Py, G., Koralsztein, J.-P., & Mercier, J. (2003). The concept of maximal lactate steady state: A bridge between biochemistry, physiology and sport science. Sports Medicine, 33(6), 407–426.
Brooks, G. A. (1991). Current concepts in lactate exchange. Medicine and Science in Sports and Exercise, 23(8), 895–906.
Brooks, G. A., Fahey, T. D., & White, T. P. Baldwin, K. M. (2000). Exercise physiology: Human bioenergetics and its applications (3rd ed.). New York, NY: McGraw-Hill Companies.
Dovalil, J., Choutka, M., Svoboda, B., Hošek, V., Perič, T., Potměšil, J., Vránová, J., & Bunc, V. (2009). Výkon a trénink ve sportu. Praha: Olympia.
Gastin, J. P. (2007). Quantification of anaerobic capacity. Scandinavian Journal of Medicine and Science in Sports, 4(2), 91–112.
Grasgruber, P., & Cacek, J. (2008). Sportovní geny. Brno: Computer Press.
Hamar, D., & Lipková, J. (2001). Fyziológia telesných cvičení. Bratislava: Univerzita Komenského.
Komi, P. V. (Ed.). (2003). Strength & power in sport (2nd ed.). Oxford: Blackwell Scientific Publications.
Máček, M., & Máčková, J. (1995). Fyziologie tělesných cvičení. Praha: SRZTV.
Máček, M., & Radvanský, J. (2011). Fyziologie a klinické aspekty pohybové aktivity. Praha: Galén.
Máček, M., & Vávra, J. (1988). Fyziologie a patofyziologie tělesné zátěže. Praha: Avicenum.
Rokyta, R. et al. (2000). Fyziologie. Praha: ISV.
Stejskal, P. (2007). Konec tradičního pojetí energetických zón? In K. Hůlka & F. Neuls (Eds.), Sborník referátu z 6. mezinárodního vědeckého semináře Efekty pohybového zatížení v edukačním prostředí tělesné výchovy a sportu (pp. 9–10). Olomouc: Univerzita Palackého.
Trojan, S. (2003). Lékařská fyziologie. Praha: Grada Publishing.